Understanding the error of Multispecies Biodiversity Indicators: A simulated example¶
Challenge and Methodological Approach Summary¶
A common benchmark for monitoring progress towards species abundance targets is the Multispecies Biodiversity Indicator (MSI). MSIs have been defined in various ways, but to us the term is best described as an estimate of the average rate of change in abundance, relative to some reference time, across a predefined set of species and geographic area. A prominent example, which was recently reinstated as a component indicator for monitoring progress towards the Global Biodiversity Framework (GBF), is the Living Planet Index (LPI). According to its website, the LPI measures the average rate of change in population sizes of native vertebrate species globally. Other examples include the EU’s grassland butterfly index and England’s all species index, which will be used to measure progress towards the respective governments’ legal commitments.
MSIs have nominal spatial and taxonomic extents that should, in theory, align with the relevant species abundance target. Spatial extents might be defined in terms of, say, a country or administrative unit (or even globally in the case of the LPI), and they can be divided conceptually into areal units or sites (e.g. grid squares on a map). Taxonomic extents are usually defined in terms of a set of species. In statistical parlance, the complete set of sites and species to which an MSI nominally pertains is known as the target population or simply the population (not to be confused with the ecological concept of a population).
Given the limited spatial and taxonomic coverage of biodiversity data, it is likely that the set of sites and species for which abundance data are available will differ from the population. It follows that the MSI obtained using the data in hand is likely to differ from the one that would have been obtained had all species and locations in the population been sampled. To use more statistical language, the sample-based MSI is known as the estimator, and the population MSI is the target parameter or estimand. Since it is the estimand that is of interest, the hope is that the proportional difference between it and the estimator, the estimation error, is small.
The ratio of the estimator to the estimand gives the proportional, or relative, estimation error. Colleagues and I decomposed the relative error of MSIs algebraically into a cross-species component reflecting the impact of non-sampled species and a within-species component reflecting the impact of non-sampled sites. Define the following quantities:
: the observed MSI at time given sampled species and sites.
: the true MSI at time given all species and sites in the target population.
: the set of species included in the sample at time ; is its size.
: the total number of species in the population at time .
: the population growth rate in mean occupancy at time relative to time 1.
: the true mean occupancy of species across all sites in the population at time .
: the sample mean occupancy of species across sampled sites at time .
Then, the full decomposition is
Notice that the error essentially boils down to the differences between sample and population means. The cross-species component is literally the difference between the sample and population mean growth rates across species. The numerators within the logs in the within-species error component represent the differences between the sample and population mean occupancy in time-periods t and 1 (i.e. the baseline year). In most realistic circumstances [1], reducing the differences between these sample and population means will reduce the cross- and within-species error components and thus the total relative error.
Meng (2018) re-expressed the differences between sample and population means in terms of three conceptually interesting quantities, and we can exploit his expression to further decompose the cross- and within-species error components. Let denote the sample mean of an arbitrary variable and be its population mean. Meng showed that
where is the data defect correlation (i.e., the correlation between the variable and the sample inclusion indicator ), is the problem difficulty (i.e. the population standard deviation of ), and is the sampling fraction, where is the sample size and is the population size. By substituting for and , we can apply Meng’s decomposition to the cross- and within-species error components of MSIs, respectively. Doing so gives us a cross-species variant of each of the three quantities and a within-species variant for every species (e.g. a within-species data defect correlation for species j, etc.).
Meng’s decomposition is remarkable, because it shows that three, and only three, factors affect the difference between a sample and population mean. The data defect correlation determines the sign and to some extent the magnitude of the difference (error). The problem difficulty and sampling fraction further scale the magnitude of the error. Of course, if one knew the values of the data defect correlation, the problem difficulty and the sampling fraction, then they could calculate the error of the sample mean. Unfortunately, it is only the sampling fraction that is measurable (assuming a well-defined population); the other two quantities must be assessed qualitatively or otherwise approximated.
In this document, I demonstrate how one might approximate the values of the cross- and within-species data defect correlations and problem difficulties. Rather than using real data, I simulate a virtual community of species in a virtual landscape comprising 1000 sites. Non-zero data defect correlations and problem difficulties are introduced by design.
Simulated example¶
First load in the required packages.
# Load required libraries
library(dplyr)
library(ggplot2)
library(tidyr)Create a virtual universe¶
The universe comprises two years and a landscape made up of 1000 sites. Each site is characterised by a variable , which varies slightly between years. The probability that each site was sampled in a given year is a function of . Since there is an interaction between year and , the effect of varies slightly between years.
set.seed(301)
n_sites <- 1000
years <- c(1, 2)
nspecies <- 20 # increased to 20 species
# Site covariates for occupancy and inclusion
z_base <- rnorm(n_sites)
z_year2 <- z_base + rnorm(n_sites, mean = 0, sd = 0.3)
# Expand site-year grid
site_year_df <- expand.grid(site = 1:n_sites, year = years) %>%
arrange(site, year) %>%
mutate(
z = ifelse(year == 1, z_base[site], z_year2[site])
)
# Generate sample inclusion once for all site-years
gamma_0 <- -1.2
gamma_year <- 0.5
gamma_z <- 0.7
gamma_int <- 0.5
site_year_df <- site_year_df %>%
mutate(
logit_pi = gamma_0 + gamma_year * year + gamma_z * z + gamma_int * year * z,
pi = plogis(logit_pi),
I = rbinom(n = n(), size = 1, prob = pi)
) %>%
select(site, year, z, I)Function to simulate species occupancy¶
The probability that any one site is occupied by a focal species in a given year is also given as a function of and year. Since determines both species occupancy probabilities and sample inclusion probabilities, it induces a dependence between the two (i.e. a non-zero within-species data defect correlation).
simulate_occupancy <- function(alpha, beta_year, beta_z, z_vals) {
z <- z_vals$z
n <- nrow(z_vals)
logit_phi <- alpha + beta_year * z_vals$year + beta_z * z
phi <- plogis(logit_phi)
Y <- rbinom(n = n, size = 1, prob = phi)
data.frame(
site = z_vals$site,
year = z_vals$year,
z = z,
Y = Y,
phi = phi
)
}Simulate a virtual community of species¶
Now we will use the function described above to simulate a virtual community of 20 species. For simplicity, we will randomly draw parameters for the function that gives the occupancy probabilities from uniform distributions whose range should ensure a mix of increasing and decreasing trends.
species_params <- lapply(1:nspecies, function(i) {
list(
alpha = runif(1, -1.5, 0.5), # baseline occupancy
beta_year = runif(1, -0.8, 0.4), # some declining, some increasing
beta_z = runif(1, -0.5, 1) # positive covariate effect
)
})
names(species_params) <- paste0("sp", 1:nspecies)
# Simulate all species
species_data_list <- lapply(names(species_params), function(sp_name) {
p <- species_params[[sp_name]]
occ_data <- simulate_occupancy(
alpha = p$alpha,
beta_year = p$beta_year,
beta_z = p$beta_z,
z_vals = site_year_df
)
occ_data$species <- sp_name
return(occ_data)
})
# Combine species and add shared inclusion
all_species <- bind_rows(species_data_list) %>%
left_join(site_year_df[, c("site", "year", "I")], by = c("site", "year"))
# Filter sampled sites
sampled_data <- all_species %>% filter(I == 1)Calculate growth rates¶
The next step is to convert each species’ values of occupancy in years one and two into a growth rate (their ratio). Growth rates are how population trends are usually formulated in MSIs.
compute_growth <- function(df) {
df %>%
group_by(species, year) %>%
summarise(mean_occ = mean(Y), .groups = "drop") %>%
pivot_wider(names_from = year, values_from = mean_occ, names_prefix = "year_") %>%
mutate(growth_rate = year_2 / year_1)
}
growth_all <- compute_growth(all_species) %>%
mutate(dataset = "all")
growth_sampled <- compute_growth(sampled_data) %>%
mutate(dataset = "sampled")
growth_summary <- bind_rows(growth_all, growth_sampled) %>%
select(species, dataset, year_1, year_2, growth_rate)Define sample inclusion for species¶
We will assume that species’ sample inclusion probabilities depend on their values of (i.e. their sensitivity to ), which can be considered a trait.
# Extract trait (beta_z) per species from species_params
traits_df <- data.frame(
species = names(species_params),
trait = sapply(species_params, function(p) p$beta_z)
)
# Define parameters for trait-based inclusion
gamma_0 <- -1 # baseline inclusion probability on logit scale
gamma_1 <- 2 # strength of trait effect on inclusion
# Compute inclusion probabilities and simulate Bernoulli draws
traits_df <- traits_df %>%
mutate(
logit_pi = gamma_0 + gamma_1 * trait,
pi = plogis(logit_pi),
included_in_sample = rbinom(n = n(), size = 1, prob = pi) == 1
)
# Remove any earlier inclusion or trait columns before joining
growth_summary <- growth_summary %>%
select(-starts_with("included_in_sample"), -starts_with("trait")) %>%
left_join(traits_df, by = "species") %>%
mutate(
log_growth_rate = log(growth_rate)
)
# View updated summary
print(growth_summary)# A tibble: 40 × 10
species dataset year_1 year_2 growth_rate trait logit_pi pi
<chr> <chr> <dbl> <dbl> <dbl> <dbl> <dbl> <dbl>
1 sp1 all 0.268 0.195 0.728 0.665 0.330 0.582
2 sp10 all 0.451 0.553 1.23 0.573 0.145 0.536
3 sp11 all 0.154 0.12 0.779 0.297 -0.406 0.400
4 sp12 all 0.364 0.241 0.662 0.607 0.213 0.553
5 sp13 all 0.311 0.176 0.566 -0.304 -1.61 0.167
6 sp14 all 0.156 0.083 0.532 0.610 0.220 0.555
7 sp15 all 0.229 0.184 0.803 0.696 0.392 0.597
8 sp16 all 0.341 0.213 0.625 0.588 0.176 0.544
9 sp17 all 0.43 0.294 0.684 0.588 0.177 0.544
10 sp18 all 0.58 0.609 1.05 -0.314 -1.63 0.164
# ℹ 30 more rows
# ℹ 2 more variables: included_in_sample <lgl>, log_growth_rate <dbl>Validating Meng’s identity: an aside¶
Now we will pause to check Meng’s identity is exact for the difference between the sample and population mean log growth rates across species. Barring some rounding error, it is correct.
growth_top <- growth_summary %>%
filter(dataset == "all") %>%
mutate(inclusion_numeric = as.numeric(included_in_sample))
y_sample_mean <- mean(growth_top$log_growth_rate[growth_top$included_in_sample])
y_pop_mean <- mean(growth_top$log_growth_rate)
diff_mean <- y_sample_mean - y_pop_mean
cor_inclusion_y <- cor(growth_top$inclusion_numeric, growth_top$log_growth_rate)
sd_y <- sd(growth_top$log_growth_rate)
f <- mean(growth_top$inclusion_numeric)
cor_identity <- cor_inclusion_y * sd_y * sqrt((1 - f) / f)
result_check <- data.frame(
mean_diff = diff_mean,
correlation_identity = cor_identity
)
print(result_check) mean_diff correlation_identity
1 0.07211732 0.07399081Assessing cross-species error¶
Assume that the analyst has data on the trait for every species, but it is measured with (random) error. One way to diagnose a non-zero cross-species data defect correlation is to compare the sample and population distributions of the measured trait values. To estimate the cross-species problem difficulty, i.e. the population standard deviation of the growth rates across species, one can use the sample standard deviation. The sample standard deviation tends be smaller than its population counterpart but can serve as a lower bound. The cross-species sampling fraction, which is to say the fraction of species in the population that were sampled, is known for a well-defined population.
# Create X = trait + epsilon (epsilon ~ N(0, 0.05))
growth_summary <- growth_summary %>%
mutate(
X = trait + rnorm(n(), mean = 0, sd = 0.05)
)
# Split dataset to match analyst's access
# Only dataset == "all" has one row per species
growth_all_only <- growth_summary %>%
filter(dataset == "all")
# The analyst only sees:
# - log_growth_rate
# - X
# - inclusion status (but not the full population)
# So they might compare included vs. not-included X values to assess correlation
# 1. Analyst tries to assess whether inclusion depends on X
# (proxy for trait that determines sampling bias)
cor_inclusion_X <- cor(growth_all_only$included_in_sample, growth_all_only$X)
# Compare the distribution of X for included vs. not
ggplot(growth_all_only, aes(x = X, fill = included_in_sample)) +
geom_density(alpha = 0.5) +
labs(title = "Distribution of Trait Proxy X by Sample Inclusion",
x = "Trait Proxy (X)", fill = "Included in Sample") +
theme_minimal()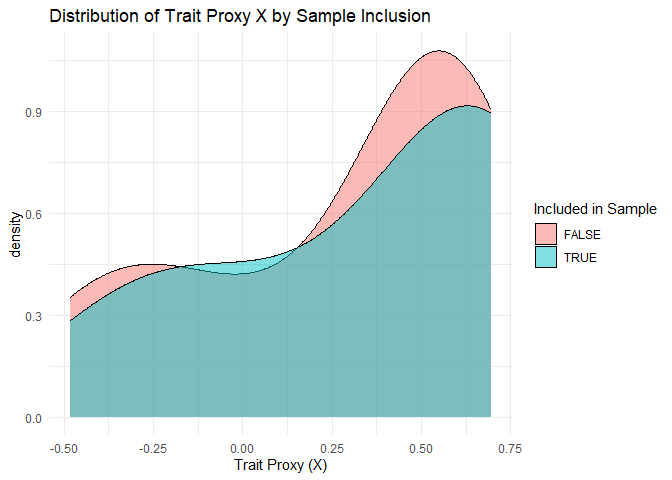
what value
1 cor(inclusion, growth rate) 0.2190484
2 cor(inclusion, X) 0.0852794
3 sd(log growth) - sample 0.1526563
4 sd(log growth) - true 0.2478649Assessing within-species error¶
The within-species data defect correlation and problem difficulty can be assessed in the same way as their cross-species counterparts. We will assume that data on is available, but measured with random error, and compare its sample and population distributions. I have simulated the data in such a way that is the only explanatory variable affecting species’ occupancy probabilities in a given year, and every species has a non-zero probability of being observed given that it occupies a sampled site. These two simplifications make assessing the within-species data defect correlations much simpler, because the set of sampled sites does not differ between species, and we need only consider rather than a larger set of explanatory variables. Hence, we can simply compare the distribution of the measured proxy for at the set of sampled sites, which does not vary between species, with its population distribution. Like the cross-species problem difficulty, we can obtain lower bounds for the within-species problem difficulties using their sample analogues. The within-species sampling fractions are directly measurable from the available data.
rho <- 0.5
n <- n_sites
# Generate epsilon
epsilon_base <- rnorm(n)
epsilon_year2 <- rnorm(n)
# x = rho * z + sqrt(1 - rho^2) * epsilon
x_base <- rho * z_base + sqrt(1 - rho^2) * epsilon_base
x_year2 <- rho * z_year2 + sqrt(1 - rho^2) * epsilon_year2
# Add to site_year_df
site_year_df$x <- ifelse(site_year_df$year == 1, x_base[site_year_df$site], x_year2[site_year_df$site])
# Prepare data
plot_df <- site_year_df %>%
mutate(sampled = ifelse(I == 1, "Sampled", "Population"))
# Faceted density plot: one panel per year
ggplot(plot_df, aes(x = x, fill = sampled)) +
geom_density(alpha = 0.5) +
facet_wrap(~year) +
scale_fill_manual(values = c("grey70", "dodgerblue")) +
labs(
title = "Distribution of Covariate Proxy x: Sampled vs. Population",
x = "x (proxy for z)",
fill = ""
) +
theme_minimal()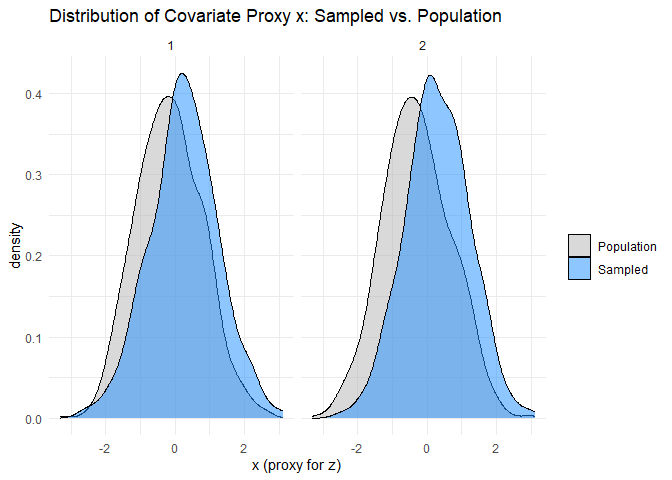
# A tibble: 40 × 3
species year ddc
<chr> <dbl> <dbl>
1 sp1 1 0.211
2 sp1 2 0.0765
3 sp10 1 0.112
4 sp10 2 0.146
5 sp11 1 0.00366
6 sp11 2 0.0104
7 sp12 1 0.150
8 sp12 2 0.121
9 sp13 1 -0.0530
10 sp13 2 -0.0323
# ℹ 30 more rows# A tibble: 40 × 4
species year pop_sd sample_sd
<chr> <dbl> <dbl> <dbl>
1 sp1 1 0.443 0.489
2 sp1 2 0.396 0.420
3 sp10 1 0.498 0.500
4 sp10 2 0.497 0.483
5 sp11 1 0.361 0.363
6 sp11 2 0.325 0.330
7 sp12 1 0.481 0.499
8 sp12 2 0.428 0.458
9 sp13 1 0.463 0.449
10 sp13 2 0.381 0.369
# ℹ 30 more rowsIn theory, the differences between sample and population mean occupancy for a given species need not be small to render a small within-species error component, so long as they have a similar sign and magnitude in both periods. How the analyst would know that they are in this situation is unclear, however, so a better strategy is to aim for zero error in both periods.